O termo fotossíntese significa síntese pela luz, sendo o processo pelo qual plantas, algas e algumas bactérias utilizam a energia luminosa para produzir matéria orgânica. A fotossíntese é o principal meio de produção de energia dos seres autotróficos. Esse processo geralmente utiliza gás carbônico (CO2) e água (H2O) para a produção de matéria orgânica na forma de glicídios, a qual servirá de alimento para o organismo, liberando também gás oxigênio (O2) para a atmosfera no processo. Praticamente todo o oxigênio que compõe a atmosfera atual da Terra é resultado da fotossíntese.
Dessa forma, a equação geral da fotossíntese é:
12 H2O + 6 CO2 → 6 O2 + C6H12O6 + 6 H2O
Conteúdo deste artigo
- Estrutura das células fotossintetizantes
- Reações da fotossíntese
- Fotofosforilação
- Fotofosforilação cíclica
- Fotofosforilação acíclica
- Fotólise da água
- Ciclo das pentoses ou ciclo de Calvin-Benson
- Primeira etapa do ciclo das pentoses
- Segunda etapa do ciclo das pentoses
- Terceira etapa do ciclo das pentoses
- O destino dos glicídios
- Aula sobre fotossíntese
- Exercícios e questões de vestibulares
Estrutura das células fotossintetizantes
Nas células, a fotossíntese ocorre em organelas chamadas de cloroplastos, a qual contém pigmentos responsáveis pela absorção da energia luminosa. O pigmento fotossintetizante mais conhecido é a clorofila, mas alguns organismos utilizam carotenoides e ficobilinas.
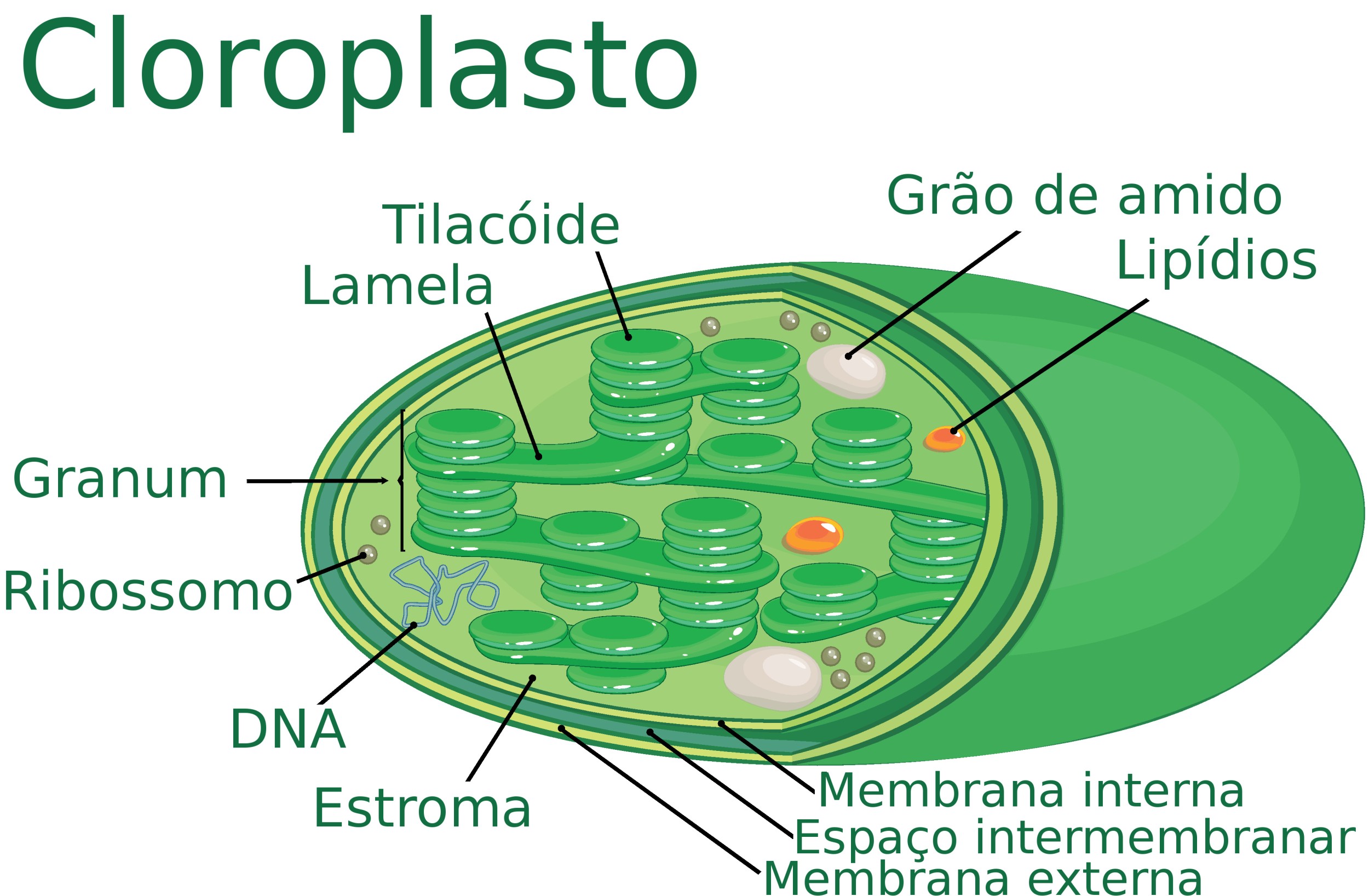
Estrutura básica de um cloroplasto, organela celular onde ocorre a Fotossíntese. Ilustração: Kazakova Maryia / Shutterstock (adaptado)
As membranas internas dos cloroplastos, chamadas de tilacóides, contêm complexos chamados de fotossistemas. Esses fotossistemas são compostos pelos pigmentos fotossintetizantes juntamente com proteínas e moléculas transportadoras de elétrons. Eles são divididos em fotossistema I e fotossistema II de acordo com o comprimento de onda considerado ótimo para a absorção.
Os dois fotossistemas funcionam de forma independente, mas são ligados por uma cadeia de moléculas transportadoras de elétrons. Os fotossistemas se organizam ainda em dois complexos: complexo antena e centro de reação. O complexo antena é formado por moléculas de pigmento que captam a energia da luz e a transfere até chegar ao centro de reação. O centro de reação é o local onde a energia luminosa vai ser convertida em energia química.
Reações da fotossíntese
A fotossíntese é composta por uma série de reações químicas. Estas reações são divididas em duas fases na fotossíntese: a fase clara ou fotoquímica e a fase escura ou puramente química.
A fase clara ocorre durante o dia, pois depende da presença de luz para acontecer. Essa fase ainda pode ser dividida em dois processos: a fotofosforilação e a fotólise da água. Já a fase escura, não depende da luz para ocorrer, sendo composta pelo processo chamado de ciclo das pentoses ou ciclo de Calvin-Benson.
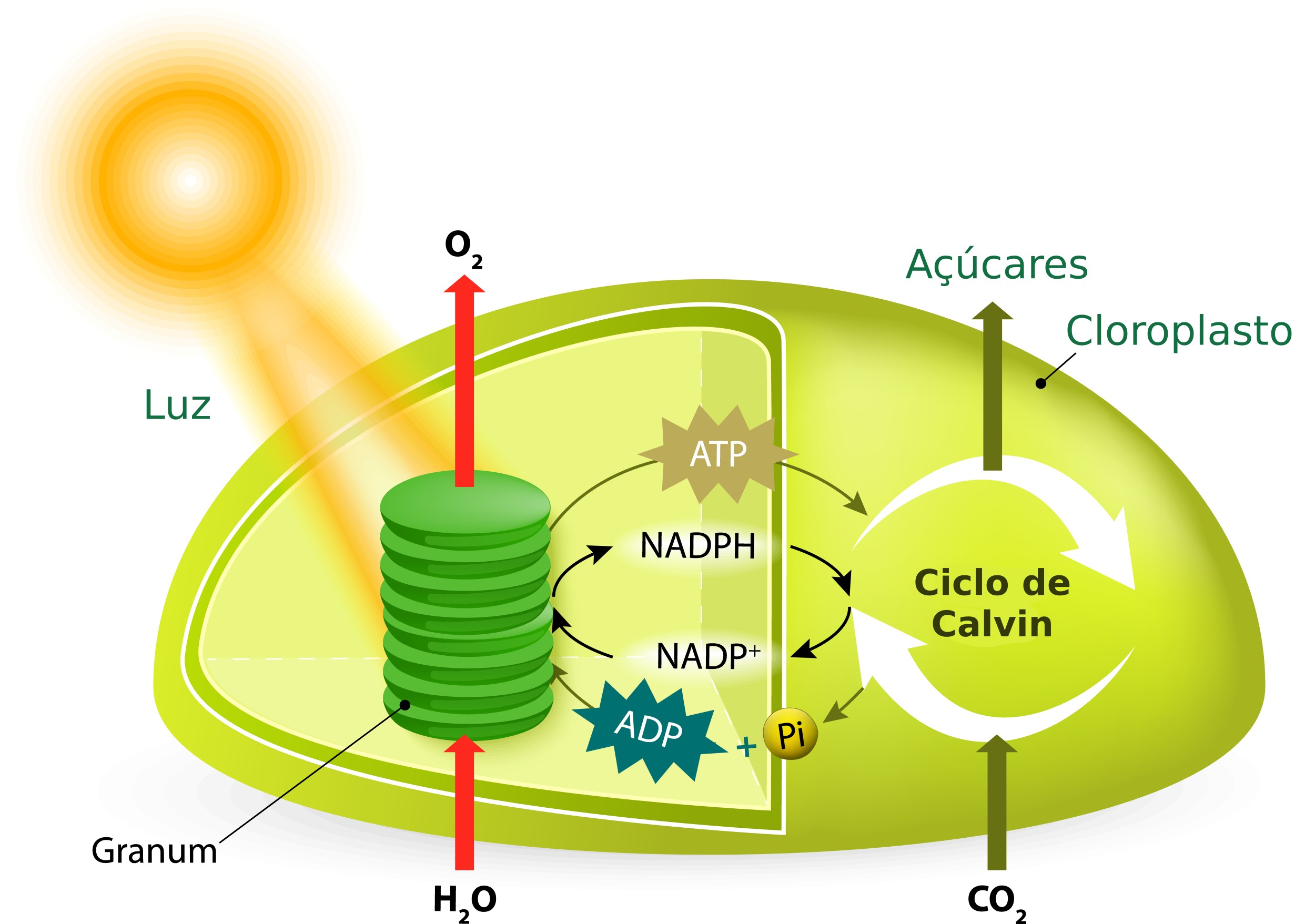
Reações do processo de fotossíntese. Ilustração: Designua / Shutterstock.com (adaptado)
Fotofosforilação
A fotofosforilação é o processo de produção de energia na forma de adenosina-trifosfato (ATP) a partir da energia luminosa. A energia luminosa que incide sobre os organismos fotossintetizantes causa uma excitação dos elétrons de uma ou mais moléculas de clorofila do complexo antena. As moléculas de clorofila excitadas transferem a energia, mas não o elétron, para moléculas de clorofila vizinhas do complexo antena, excitando-as também. Essa transferência vai ocorrendo até chegar ao centro de reação do fotossistema.
Quando uma molécula de clorofila do centro de reação recebe a energia, seus elétrons são excitados de forma a atingir um nível superior de energia e serem transferidos para uma molécula transportadora de elétrons. A fotofosforilação pode ser de dois tipos: fotofosforilação cíclica e acíclica.
Fotofosforilação cíclica
A fotofosforilação cíclica ocorre no fotossistema I, composto basicamente por clorofila a. Ao receber a energia luminosa, um par de elétrons excitados deixa a molécula de clorofila a. A partir disso, o elétron vai sendo transferido de uma substância para a outra em uma cadeia transportadora de elétrons. Após a passagem por essa cadeia, retornam a molécula de clorofila, ocupando seu lugar e fechando a fotofosforilação cíclica.
Fotofosforilação acíclica
Já na fotofosforilação acíclica, os fotossistemas I e II trabalham em conjunto. No fotossistema II predomina a clorofila b. Durante o processo a clorofila a do fotossistema I que recebeu a energia luminosa perde um par de elétrons excitados, sendo recolhidos por uma molécula aceptora de elétrons. Esses elétrons vão sendo passados por uma cadeia transportadora de elétrons, na qual último aceptor é uma molécula chamada de NADP+ (fosfato de dinucleotídeo de nicotinamida-adenina), que se torna NADPH2 ao receber os elétrons.
No fotossistema II, a clorofila b, também excitada pela luz, perde um par de elétrons. Esse par atravessa outra cadeia transportadora de elétrons que liga os dois fotossistemas, chegando ao fotossistema I e ocupando o lugar do elétron perdido pela clorofila a. Como os elétrons que voltam para a clorofila a não são os mesmos que foram perdidos por ela, mas sim os doados pela clorofila b, essa etapa da fotossíntese é chamada de fotofosforilação acíclica.
Ao passar pela cadeia transportadora, esses elétrons liberam energia que vai ser utilizada para realizar a passagem prótons (H+) através das membranas tilacóides, passando do estroma do cloroplasto para o interior do tilacóide ou lúmem. A alta concentração de H+ acumulados no interior dos tilacóides cria uma pressão para a sua saída. A forma que esses íons encontram de sair é através de um complexo enzimático transmembrana chamado de sintetase de ATP. Esse complexo funciona como um motor molecular, que gira com a passagem de H+, unindo moléculas de ADP com fosfatos (Pi) para a produção de ATP.
Tanto esse ATP quanto o NADPH2 produzido ao final da cadeia transportadora de elétrons, vão ter um papel no ciclo das pentoses da fase escura. Vale lembrar que a molécula de clorofila b continua sem seu par de elétrons, que só vai ser reposto no processo de fotólise da água.
Fotólise da água
A molécula de clorofila que perdeu seu elétron após a excitação pela energia luminosa, é capaz de substituí-lo por elétrons extraídos de moléculas de água. Com a remoção dos seus elétrons, a molécula de água decompõe-se em prótons (H+) e átomos livres de oxigênio (O). Os prótons são liberados para dentro da membrana tilacóide, vindo a contribuir para o aumento da concentração de prótons no lúmem e a consequente geração de ATP. Já os átomos de oxigênio liberados unem-se imediatamente em pares formando moléculas de gás oxigênio (O2), que são liberadas para a atmosfera. Essa quebra provocada pela oxidação (perda de elétrons) da água em decorrência da energia luminosa é chamada então de fotólise da água.
A equação geral da fotólise da água é a seguinte:
2 H2O → 4e– + 4 H+ + O2
Ciclo das pentoses ou ciclo de Calvin-Benson
Já na fase escura da fotossíntese, ocorre o ciclo das pentoses ou ciclo de Calvin-Benson. Ele consiste em um conjunto de reações que ocorre de forma cíclica, sendo responsável pela produção de glicídios que vão servir de alimento para o organismo. Esse processo realiza a fixação do carbono atmosférico, pois utiliza moléculas de gás carbônico (CO2) como fonte de carbono para a produção dos glicídios.
O ciclo é composto por três etapas:
Primeira etapa do ciclo das pentoses
O composto inicial do ciclo é um açúcar de cinco carbonos com um grupo fosfato chamado de ribulose-1,5-bifosfato (RuBP). A partir disso, ocorre a incorporação de uma molécula de CO2 ao RuBP mediada pela enzima chamada de rubisco, o que resulta em duas moléculas de três carbonos cada, chamadas de 3-fosfatoglicerato ou ácido 3-fosfoglicérico (PGA). Dessa forma, a cada 6 moléculas de CO2 incorporadas a 6 moléculas RuBP produz-se 12 moléculas de PGA, sendo esta a quantidade necessária para a realização do ciclo completo e produção de 1 molécula de glicose ao final da fotossíntese.
Segunda etapa do ciclo das pentoses
Na segunda etapa, o PGA é utilizado na produção de um composto chamado de gliceraldeído 3-fosfato ou 3-fosfogliceraldeído (PGAL), de fórmula química C3H6O3. O PGAL é o principal produto do ciclo das pentoses e sua produção inclui duas reações. Na primeira delas o PGA é fosforilado, recebendo o fosfato (Pi) de uma molécula de ATP produzida na fotofosforilação da fase clara. Dessa forma, o PGA passa a ser uma molécula com dois fosfatos chamada de 1,3-bifosfoglicerato e o ATP volta à condição de ADP. A partir disso, ocorre a redução do 1,3-bifosfoglicerato pelo NADPH2 também produzido pela fotofosforilação. Nessa reação de redução o 1,3-bifosfoglicerato tem um dos seus fosfatos removido gerando o PGAL, enquanto o NADPH2 volta à condição de NADP+.
Terceira etapa do ciclo das pentoses
Na terceira etapa, das 12 moléculas de PGAL produzidas, 10 são utilizadas para a regeneração das 6 moléculas de RuBP necessárias para o ciclo se iniciar novamente. Como foi visto, o ciclo das pentoses não forma diretamente a glicose (C6H12O6), mas sim o glicídio chamado de gliceraldeído 3-fosfato (PGAL). As 2 moléculas de PGAL que não são utilizadas para regenerar a RuBP, saem do ciclo, podendo ser transportadas para o citoplasma da célula. A partir dessas duas moléculas de PGAL pode ser produzida uma molécula de glicose.
Assim, desconsiderando a formação posterior de glicose, a produção mais direta de glicídios na fotossíntese é representada pela seguinte fórmula:
3 CO2 + 6 H2O → C3H6O3 + 3 O2 + 3 H2O
O destino dos glicídios
Embora seja comumente representada como o principal carboidrato produzido na fotossíntese, a glicose livre é gerada em baixa quantidade nas células fotossintetizantes. Na verdade, a maior parte das moléculas de PGAL que saem para o citoplasma forma o açúcar sacarose, composto por uma molécula de glicose e uma de frutose.
Parte dos glicídios da fotossíntese é utilizada pelas mitocôndrias na respiração celular, produzindo energia para o organismo realizar suas funções vitais. Outra parte pode vir a integrar a biomassa do ser fotossintetizante através da produção de diversas substâncias orgânicas como aminoácidos, gorduras e celulose. A glicose ainda pode ser convertida em amido e ser armazenada em células especiais do caule e da raiz compondo uma reserva energética para o organismo.
Aula sobre fotossíntese
Assista abaixo uma aula do Canal Futura sobre Fotossíntese:
Referências:
Amabis, J. M. & Martho, G. R. 2006. Fundamentos da Biologia Moderna: Volume único. 4ª Ed. Editora Moderna: São Paulo, 839 p.
Raven, P. H.; Evert, R. F.; Eichorn, S. E. 2007. Biologia vegetal. 7a ed. Editora Guanabara Koogan: Rio de Janeiro, 830 p.
Texto originalmente publicado em https://www.infoescola.com/biologia/fotossintese/
